






啮齿类动物的雌性生殖生理
来源:未知 |发布时间:2018-06-14 09:59|点击:次
啮齿类动物的雌性生殖生理
卵巢是雌性啮齿类动物的主要生殖器官,能够产生并排出成熟的卵子,分泌雌激素、孕激素等卵巢激素,参与生殖系统的协调活动和影响机体其他组织功能。卵子发生是指雌配子的形成、发育和成熟。包括卵原细胞的增殖、卵母细胞的生长和卵母细胞的成熟等几个阶段。卵子的成熟分裂分为前期(包括细线期、偶线期、粗线期、双线期和终变期)、中期和末期。第1次成熟分裂后初级卵母细胞分裂为次级卵母细胞和第1极体,第1极体以后还可分裂为两个极体。而第2次成熟分裂使次级卵母细胞分裂为卵细胞和第2极体。第2次成熟分裂在受精过程中完成,成熟卵子的染色体数目只有原来的一半。大多数啮齿类动物在排卵时卵母细胞已经完成第1次成熟分裂。排卵后,次级卵泡开始第2次成熟分裂,到受精时产生第2极体完成第2次成熟分裂。
母鼠在排卵前释放的促性腺激素可促使卵泡成熟分裂,有利于卵母细胞的成熟。将卵母细胞从卵泡中取出培养,即使没有促性腺激素也会自发成熟,因而推测卵泡可能产生某些抑制因子阻遏成熟分裂,而促性腺激素则能解除这种阻遏作用。卵泡发育分为原始卵泡、初级卵泡、次级卵泡、三级卵泡和成熟卵泡几个阶段。卵母细胞开始第1次成熟分裂时,周围被一层扁平的卵泡细胞包围,形成原始卵泡。原始卵泡中的初级卵母细胞生长增大,卵泡细胞分裂增殖,胞浆内出现多层颗粒细胞,发育成初级卵泡。逐渐卵黄颗粒增多,卵泡细胞增殖,初级卵母细胞与颗粒细胞之间,由颗粒细胞分泌的糖蛋白构成透明带,透明带周围的颗粒细胞呈放射状排列,称放射冠。放射冠细胞的突起可穿进透明带与卵母细胞接触并提供营养。次级卵泡中卵泡细胞逐渐分离,形成腔隙,其中充满由卵泡细胞分泌的卵泡液,随着卵泡腔的扩大和卵泡液的增多,卵母细胞被推向一边,并被包裹在卵泡细胞团中形成卵丘,颗粒层细胞贴在卵泡腔周围形成颗粒层,这时卵泡已发育成三级卵泡。成熟卵泡的卵泡膜分为外膜和内膜两层,内膜细胞参与雌激素的合成。卵泡内的初级卵母细胞中断的第1次成熟分裂开始恢复。
在卵泡发育过程中卵母细胞的生长、卵泡细胞的增殖、卵泡腔的形成和脱离原始卵泡库等都受到垂体促性腺激素的调节。切除脑垂体后大鼠、小鼠卵泡的早期生长过程仍然可以继续进行,原始卵泡的发育不依靠垂体激素。在卵泡内有4层颗粒细胞,特别是卵泡内膜形成后,卵泡的生长依靠垂体激素。而仓鼠在摘除垂体后卵巢上小腔前卵泡数减少,用外源性促性腺激素可以消除这种影响,说明垂体促性腺激素对原始卵泡的生长是有促进作用的。
卵泡中雌激素含量的减少和雄激素含量的增加,可能引起闭锁卵泡基膜通透性发生改变,使得一些原本排除在外的血浆蛋白成分进入卵泡,甚至一些血浆成分如IcG、IJM和血液有形成分也可进入卵泡,造成颗粒细胞的溶解和固缩。输卵管由呈漏斗状的伞靠近卵巢向腹腔开口的漏斗部、较远侧的扩大部壶腹、较近侧连接输卵管与子宫腔的底部等几部分组成。输卵管的肌肉系统、上皮纤毛细胞和分泌细胞等结构能够适应多种功能,在排卵时,兔和鼠的伞部将卵巢包围成一个卵巢囊。卵巢和输卵管卵巢韧带、输卵管系膜以及输卵肌肉间歇性收缩,排出的卵子在卵巢囊内做不定向的移动,卵子与伞部的纤毛密切接触。由于纤毛朝一个方向摆动,卵子就能很快地通过伞部表面进入输卵管。而有些啮齿动物如小鼠、大鼠和田鼠伞部只有小部分与卵巢发生接触。输卵管系膜的收缩受甾体激素的调节。
卵巢主要分泌雌激素、孕激素和少量的雄激素、抑制素。在排卵前由卵泡颗粒细胞层分泌雌激素;排卵后,由黄体分泌孕激素和雌激素。雌二醇,雌酮和雌三醇是卵巢分泌的主要雌激素。卵子起源于卵巢的卵原细胞。卵巢内的卵细胞的生成和卵泡的发育同步。原始的卵泡由一个初级卵母细胞和包围它的颗粒细胞组成。胚胎时的卵囊移至卵巢内,分化为卵原细胞,经过多次有丝分裂,成为初级卵母细胞,这个过程叫卵子的发生( oogenesis)或叫为增殖期(图2-14)。初级卵母细胞外面包一层卵泡细胞成为初级卵泡,该阶段可持续很长时间,直到雌性动物达到性成熟,卵泡才进入生长和成熟期。大多数动物的卵母细胞仅停留在第1次成熟分裂前期,直至排卵前。卵母细胞发育的同时,周围的卵泡细胞由一层变为多层,细胞增大,细胞质内含有颗粒,成为颗粒细胞层。卵泡周围的结缔组织及间质细胞相继形成卵膜及其内侧的卵泡膜细胞层。颗粒细胞层、卵泡膜细胞层都具有内分泌功能。卵泡成熟以后,卵母细胞被排出,卵泡膜塌陷,其颗粒细胞肥大,与卵膜内细胞层共同转化为黄体细胞,并构成黄体,黄体也具有内分泌功能。

啮齿类动物的雌激素促进输卵管上皮增生,分泌与运动加强;促进子宫发育,内膜增生,分泌大量清亮稀薄液体,其中的黏蛋白沿子宫颈纵行排列,有利于精子穿行。
雌性动物中的雄激素都是作为雌激素的前体形式存在,所以有时在卵巢中可以测到较高水平的雄激素。卵巢和睾丸一样,也能分泌抑制素,于卵泡成熟时抑制卵母细胞成熟,停留在第1次成熟分裂前期直至排卵前。
1.卵巢功能的调节
卵巢的生卵作用和内分泌功能受下丘脑一腺垂体一卵巢轴的作用。即下丘脑分泌促性腺释放激素(GnRH)可促进腺垂体合成和分泌促性腺激素(GtH);促性腺激素引起性腺合成和分泌性激素,并影响到性腺中卵泡发育、成熟及排卵。性腺分泌的性激素对腺垂体和下丘脑有反馈性作用(图2-15)。现已证明:啮齿类动物在原始卵泡及初级卵泡的早期发育阶段基本上不受垂体的调控,主要取决于卵泡内部因子。如GH、胰岛素或胰岛素样生长因子,可刺激颗粒细胞增生;颗粒细胞的分泌物又可促进卵泡膜的形成。初级卵泡发育期,卵泡中可能有一种促进促卵泡激素(FSH)分泌的蛋白( FSH-releasing protein,FRP)能使FSH的分泌增加。

性成熟是需要经历若干时间的发展过程。性成熟过程的开始阶段叫初情期( puberty)。性成熟的各种变化是由睾酮或雌激素分泌增多而实现的。而性激素合成和释放又与下丘脑垂体系统有复杂的相互作用。不同种的鼠达到性成熟的年龄各不相同,如鼢鼠要经过1年后才能达到性成熟,而田鼠则可在2个月达到性成熟。常年繁殖的动物,如家鼠,初情期多在体重达到成年动物的40%~60%或生长速度下降的初期。季节性繁殖动物的初情期受出生期的影响很大。
啮齿类动物的繁殖受光照、温度和食物来源等环境因素的影响,一般都在最适宜妊娠和幼子生活的植物生长季节繁殖;而家鼠由于环境因素和食物来源比较稳定,经过长期进化,其繁殖季节逐渐延长,甚至可以全年繁殖。啮齿类动物达到性成熟后,雌鼠会在一年繁殖期中有规律地多次动情,而雄鼠则会在整个繁殖期不断形成精子,因而出现生殖季节,如家鼠、家兔等。常年繁殖并不意味全年的繁殖活动毫无变化。大多数常年繁殖鼠种的生殖活动,在不同季节表现出有规律的高峰期和低潮期。
典型的季节性繁殖啮齿类动物每年只出现一个或两个繁殖季节。在繁殖季节内,雌鼠能一次或多次动情,雄鼠能不断形成精子。但在休情期内,卵巢或睾丸都不同程度地萎缩,甚至退化到幼鼠那样的程度,两性配子的形成完全停止。在休情期内,睾丸常缩回腹腔,各种依赖性激素控制的附性器官也都退化。
性周期,雌性动物在性成熟后,卵巢在神经和体液的调节下,出现周期性的卵泡成熟期和排卵期伴随着每次卵泡的成熟和排卵,整个机体特别是生殖器官,发生系列的形态和机能的变化,同时动物还出现周期性的性反射和性行为过程,称为动情周期( estrouscycle)或性周期( sexual cycle)。这种周期性的性活动过程除了妊娠期之外,一直延续到性机能停止的年龄为止。3种鼠类的动情期见表2-4。

动情周期是一系列逐渐变化的复杂生理过程,难以严格地加以区分,通常将其分为动情前期、动情期、动情后期和间情期。
动情前期( proestms)是性周期的准备阶段和性活动开始的时期。这时动物处于安静状态,没有交配欲的表现,但生殖器官却发生一系列变化,如卵巢内卵泡迅速生长,并达到成熟阶段;卵泡液体积增加,雌激素分泌并释放到血液中,刺激输卵管内壁的细胞生长,纤毛增多;子宫角蠕动加强,子宫内膜的血管分布大量增生;阴道上皮细胞增生加厚,整个生殖道腺体活动力增强。
动情期( estrus)是性周期的高潮期。这时动物出现兴奋不安,食欲减退,时常鸣叫和爬跨其他个体,并有交配欲。生殖器官的变化是:大多数动物出现卵巢中的成熟卵泡破裂和排卵,子宫黏膜血管大量增生、腺体分泌、子宫颈口张开、出现子宫蠕动和水肿,输卵管岀现蠕动,阴唇黏膜肿胀,有些啮齿动物从阴道流岀黏液。动情期啮齿类动物的各种表现和生殖器官的形态和机能的变化,其主要原因是由于卵泡所分泌的雌激素的作用所产生的。在一般情况下,啮齿类动物动情的同时往往引起排卵,但是,动情和排卵两种生理活动过程是可以分离而单独存在的。例如,切除卵巢的啮齿类动物,给予外源性雌激素仍能引起动情,但不能排卵,在生产实践中有些患有不孕的啮齿类动物,其原因之一是只有动情而不排卵。
动情后期( metaestrus)指动情结束后的一段时期。此期内雌性啮齿类动物恢复安静并拒绝交配。卵巢中形成了黄体,并分泌孕激素和雌激素。在孕激素的作用下,子宫为接受胚泡和适应胚泡的营养及附植发生一系列的变化。如果排出的卵细胞受精,动情周期就此中止,并开始进入妊娠阶段,直到分娩后才重新出现新的动情周期。如果排出的卵细胞没有受精,就过渡到间情期。
间情期( diestrus)是相对生理静止期。卵巢内的黄体开始退化,卵泡还没有开始发育。生殖道朝着恢复动情前期以前的状态退化,子宫内膜变薄,阴道上皮不角化等。
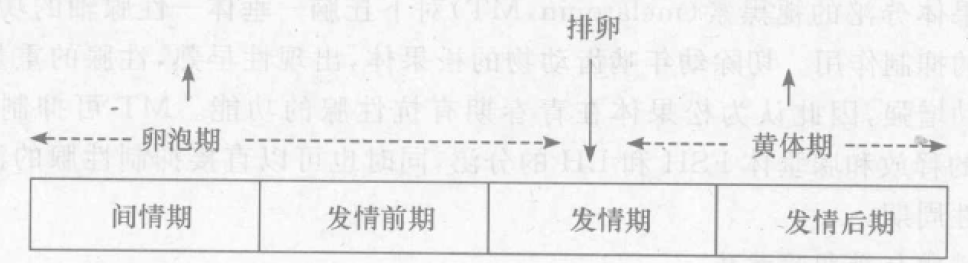
啮齿类动物的动情周期经常受到内外环境因素、营养及健康状况的影响。突然而剧烈的环境变化会造成动情周期的紊乱甚至停止。若以卵巢周期性变化特征来区分,可将性周期分为卵泡期( follicular phase)和黄体期( luteal phase),而排卵则是卵泡期和黄体期的分界线。卵巢周期和动情周期的关系为:
 3.性周期的调节
3.性周期的调节
性周期的变化经常受内外环境因素、营养及健康状况的影响,主要受神经体液调节,其中激素的调节是重要基础。外环境变化的信息以及雄性刺激由雌性啮齿类动物的嗅觉、视觉、触觉感受器所接受,经过不同途径,通过神经系统影响到下丘脑一腺垂体一卵巢轴。性周期的几个阶段实质上是雌激素与下丘脑的GnRH、腺垂体的FSH、LH的分泌活动的相互协调。
卵泡初期(即动情之初期)是雌激素积累时期。此时雌激素浓度较低,对FSH、LH的分泌具有负反馈作用,但作用十分弱;此时卵泡靠内部的自我驱动机制加快了颗粒细胞分裂、增殖,雌激素分泌功能增强。
卵泡中期(动情前期的中、后期)重要的是卵泡及血液中雌激素浓度继续上升,并出现高峰。此时,卵母细胞已进入第1次成熟分裂早期,尽管有雌激素和卵泡抑制素的抑制性负反馈作用,但由于卵泡内的糖蛋白雄激素局部正反馈作用,通过卵泡膜细胞LH受体数量的增加,最终到达雌激素分泌高峰。
卵泡后期(进入动情期,排卵前夕)LH高峰出现。这是由于雌激素高峰的出现,对下丘脑、腺垂体产生正反馈作用,GnRH脉冲式分泌量及频率都增加,腺垂体对GnRH的敏感性也增加,因而出现LH分泌高峰,LH通过刺激孕激素分泌,影响到卵内酶的活性,而触发排卵。
黄体期(动情后期)LH刺激黄体细胞分泌孕激素、雌激素,后者对下丘脑和腺垂体产生负反馈作用,使GnRH分泌下降。同时当雌激素分泌量升高时,降低了下丘脑催乳素抑制激素的释放,而引起垂体前叶催乳素释放量增加,催乳素与LH协同作用,促进和维持黄体分泌孕酮。当孕酮分泌达到一定量时,对下丘脑和腺垂体也会产生负反馈作用,抑制垂体前叶FSH的分泌,以至卵巢卵泡不再发育,抑制宠物中枢神经系统的性中枢,使雌性啮齿类动物不再表现动情。同时孕酮也作用于生殖道及子宫,使之发生有利于胚胎附植的生理变化。如果排出的卵子已受精,囊胚刺激子宫内膜形成胎盘。此时黄体则继续存在下去成为妊娠黄体;若未受精,因没有取代的促性腺激素,黄体退化,雌激素、孕激素分泌下降并终止,对下丘脑-腺垂体的负反馈作用终止,又开始下一轮的周期。
松果体分泌的褪黑素( melatonin,MT)对下丘脑一垂体一性腺轴的功能活动有明显的抑制作用。切除幼年啮齿类动物的松果体,出现性早熟,性腺的重量增加,功能活动增强,因此认为松果体在青春期有抗性腺的功能。MT可抑制下丘脑GnRH的释放和腺垂体FSH和LH的分泌,同时也可以直接抑制性腺的活动,从而影响性周期。
4.排卵与排卵后黄体
(1)排卵过程
成熟卵泡壁发生破裂,卵细胞、透明带与放射冠随同卵泡液冲出卵泡,称为排卵( ovulation)。排卵过程从卵巢表面上皮细胞的逐步溶解开始,随后细胞开始退行性变化,并释放蛋白酶进入细胞间隙,使白膜、结缔组织、卵泡外膜和内膜崩解,整个卵泡壁也发生普遍性溶解。当破口处的颗粒细胞变性破坏时,卵细胞及其放射冠随同卵泡液被排人输卵管伞或腹腔。排卵前,卵泡经历着3大变化:①卵母细胞细胞质和细胞核成熟;②卵细胞聚合力松解,颗粒细胞各自分离;③卵泡膜变薄、破裂。
(2) 排卵机制
垂体脉冲式地释放大量LH是激发排卵的必要条件,而LH的释放又受到雌激素的正反馈调控。升高的雌激素在下丘脑经2-羟化酶催化,生成儿茶酚雌醇或儿茶酚雌酮,统称儿茶酚雌激素( catecholestron),这类激素具有儿茶酚和雌激素的双重作用,一方面能减少去甲肾上腺素(NE)的降解,并增强NE对下丘脑分泌Litre的刺激作用;另一方面也能在下丘脑和腺垂体内与雌激素竞争受体,抑制雌激素对下丘脑和垂体的负反馈调节。升高的雌激素一方面刺激下丘脑GnRH脉冲式分泌频率和幅度极大地增加;另一方面也使腺垂体对GnRH的反应超常的大,使LH分泌及在血液中的水平达到顶峰。LH通过刺激孕激素的生成,增强卵泡中许多蛋白水解酶的活性。LH也直接促进卵泡大量生成蛋白水解酶。蛋白水解酶活性的增强,是卵泡破裂的主要因素。此外,孕酮、前列腺素等激素也对排卵起重要作用。它们能增强纤维蛋白分解酶原的产生,从而增加纤维蛋白分解酶的活性,使卵泡膜分解破裂;并可刺激颗粒细胞合成孕酮及卵泡黄体化。则能对溶酶体膜起破坏作用,因此,可促进卵泡顶端上皮细胞溶酶体的破裂,使上皮细胞脱落,顶端形成排卵点,所有这些都为排卵创造条件。
(3)黄体的生成和退化
成熟卵泡破裂排卵后,由于卵泡液排出,卵泡壁塌陷皱缩,从破裂的卵泡壁血管流出血液和淋巴液,并聚积于卵泡腔内形成血凝块。此后颗粒细胞增生肥大,吸收类脂物质黄素,而变成黄体细胞,构成黄体的主体部分。同时卵泡内膜分生出血管,布满于发育中的黄体。在动情周期中,雌性啮齿动物如果没有妊娠,所形成的黄体在黄体期末退化,这种黄体称为周期性黄体。黄体退化时,颗粒细胞转化的黄体细胞退化很快,体积逐渐变小,细胞的数目也显著减少,颗粒层细胞逐渐被纤维细胞所代替,黄体细胞间结缔组织侵入、增殖,最后整个黄体颜色变白称为白体( corpus albicans),残留在卵巢内。大多数啮齿类动物的白体存在到下一周期的黄体期,即此时的功能性新黄体与大部分退化的白体共存。
卵巢是雌性啮齿类动物的主要生殖器官,能够产生并排出成熟的卵子,分泌雌激素、孕激素等卵巢激素,参与生殖系统的协调活动和影响机体其他组织功能。卵子发生是指雌配子的形成、发育和成熟。包括卵原细胞的增殖、卵母细胞的生长和卵母细胞的成熟等几个阶段。卵子的成熟分裂分为前期(包括细线期、偶线期、粗线期、双线期和终变期)、中期和末期。第1次成熟分裂后初级卵母细胞分裂为次级卵母细胞和第1极体,第1极体以后还可分裂为两个极体。而第2次成熟分裂使次级卵母细胞分裂为卵细胞和第2极体。第2次成熟分裂在受精过程中完成,成熟卵子的染色体数目只有原来的一半。大多数啮齿类动物在排卵时卵母细胞已经完成第1次成熟分裂。排卵后,次级卵泡开始第2次成熟分裂,到受精时产生第2极体完成第2次成熟分裂。
母鼠在排卵前释放的促性腺激素可促使卵泡成熟分裂,有利于卵母细胞的成熟。将卵母细胞从卵泡中取出培养,即使没有促性腺激素也会自发成熟,因而推测卵泡可能产生某些抑制因子阻遏成熟分裂,而促性腺激素则能解除这种阻遏作用。卵泡发育分为原始卵泡、初级卵泡、次级卵泡、三级卵泡和成熟卵泡几个阶段。卵母细胞开始第1次成熟分裂时,周围被一层扁平的卵泡细胞包围,形成原始卵泡。原始卵泡中的初级卵母细胞生长增大,卵泡细胞分裂增殖,胞浆内出现多层颗粒细胞,发育成初级卵泡。逐渐卵黄颗粒增多,卵泡细胞增殖,初级卵母细胞与颗粒细胞之间,由颗粒细胞分泌的糖蛋白构成透明带,透明带周围的颗粒细胞呈放射状排列,称放射冠。放射冠细胞的突起可穿进透明带与卵母细胞接触并提供营养。次级卵泡中卵泡细胞逐渐分离,形成腔隙,其中充满由卵泡细胞分泌的卵泡液,随着卵泡腔的扩大和卵泡液的增多,卵母细胞被推向一边,并被包裹在卵泡细胞团中形成卵丘,颗粒层细胞贴在卵泡腔周围形成颗粒层,这时卵泡已发育成三级卵泡。成熟卵泡的卵泡膜分为外膜和内膜两层,内膜细胞参与雌激素的合成。卵泡内的初级卵母细胞中断的第1次成熟分裂开始恢复。
在卵泡发育过程中卵母细胞的生长、卵泡细胞的增殖、卵泡腔的形成和脱离原始卵泡库等都受到垂体促性腺激素的调节。切除脑垂体后大鼠、小鼠卵泡的早期生长过程仍然可以继续进行,原始卵泡的发育不依靠垂体激素。在卵泡内有4层颗粒细胞,特别是卵泡内膜形成后,卵泡的生长依靠垂体激素。而仓鼠在摘除垂体后卵巢上小腔前卵泡数减少,用外源性促性腺激素可以消除这种影响,说明垂体促性腺激素对原始卵泡的生长是有促进作用的。
卵泡中雌激素含量的减少和雄激素含量的增加,可能引起闭锁卵泡基膜通透性发生改变,使得一些原本排除在外的血浆蛋白成分进入卵泡,甚至一些血浆成分如IcG、IJM和血液有形成分也可进入卵泡,造成颗粒细胞的溶解和固缩。输卵管由呈漏斗状的伞靠近卵巢向腹腔开口的漏斗部、较远侧的扩大部壶腹、较近侧连接输卵管与子宫腔的底部等几部分组成。输卵管的肌肉系统、上皮纤毛细胞和分泌细胞等结构能够适应多种功能,在排卵时,兔和鼠的伞部将卵巢包围成一个卵巢囊。卵巢和输卵管卵巢韧带、输卵管系膜以及输卵肌肉间歇性收缩,排出的卵子在卵巢囊内做不定向的移动,卵子与伞部的纤毛密切接触。由于纤毛朝一个方向摆动,卵子就能很快地通过伞部表面进入输卵管。而有些啮齿动物如小鼠、大鼠和田鼠伞部只有小部分与卵巢发生接触。输卵管系膜的收缩受甾体激素的调节。
卵巢主要分泌雌激素、孕激素和少量的雄激素、抑制素。在排卵前由卵泡颗粒细胞层分泌雌激素;排卵后,由黄体分泌孕激素和雌激素。雌二醇,雌酮和雌三醇是卵巢分泌的主要雌激素。卵子起源于卵巢的卵原细胞。卵巢内的卵细胞的生成和卵泡的发育同步。原始的卵泡由一个初级卵母细胞和包围它的颗粒细胞组成。胚胎时的卵囊移至卵巢内,分化为卵原细胞,经过多次有丝分裂,成为初级卵母细胞,这个过程叫卵子的发生( oogenesis)或叫为增殖期(图2-14)。初级卵母细胞外面包一层卵泡细胞成为初级卵泡,该阶段可持续很长时间,直到雌性动物达到性成熟,卵泡才进入生长和成熟期。大多数动物的卵母细胞仅停留在第1次成熟分裂前期,直至排卵前。卵母细胞发育的同时,周围的卵泡细胞由一层变为多层,细胞增大,细胞质内含有颗粒,成为颗粒细胞层。卵泡周围的结缔组织及间质细胞相继形成卵膜及其内侧的卵泡膜细胞层。颗粒细胞层、卵泡膜细胞层都具有内分泌功能。卵泡成熟以后,卵母细胞被排出,卵泡膜塌陷,其颗粒细胞肥大,与卵膜内细胞层共同转化为黄体细胞,并构成黄体,黄体也具有内分泌功能。
啮齿类动物的雌激素促进输卵管上皮增生,分泌与运动加强;促进子宫发育,内膜增生,分泌大量清亮稀薄液体,其中的黏蛋白沿子宫颈纵行排列,有利于精子穿行。
雌性动物中的雄激素都是作为雌激素的前体形式存在,所以有时在卵巢中可以测到较高水平的雄激素。卵巢和睾丸一样,也能分泌抑制素,于卵泡成熟时抑制卵母细胞成熟,停留在第1次成熟分裂前期直至排卵前。
1.卵巢功能的调节
卵巢的生卵作用和内分泌功能受下丘脑一腺垂体一卵巢轴的作用。即下丘脑分泌促性腺释放激素(GnRH)可促进腺垂体合成和分泌促性腺激素(GtH);促性腺激素引起性腺合成和分泌性激素,并影响到性腺中卵泡发育、成熟及排卵。性腺分泌的性激素对腺垂体和下丘脑有反馈性作用(图2-15)。现已证明:啮齿类动物在原始卵泡及初级卵泡的早期发育阶段基本上不受垂体的调控,主要取决于卵泡内部因子。如GH、胰岛素或胰岛素样生长因子,可刺激颗粒细胞增生;颗粒细胞的分泌物又可促进卵泡膜的形成。初级卵泡发育期,卵泡中可能有一种促进促卵泡激素(FSH)分泌的蛋白( FSH-releasing protein,FRP)能使FSH的分泌增加。
2.生殖周期
啮齿类动物生长发育达到一定阶段,生殖器官和副性征的发育已经基本完成,开始具备了生殖能力,这个时期叫性成熟( sexual maturity)。在性成熟个体的性腺中开始形成成熟的两性配子(精子和卵子),表现出各种性反射,出现性的要求和配种欲望。这时雌、雄个体能交配和受精,并能完成妊娠和胚胎发育过程。性成熟是需要经历若干时间的发展过程。性成熟过程的开始阶段叫初情期( puberty)。性成熟的各种变化是由睾酮或雌激素分泌增多而实现的。而性激素合成和释放又与下丘脑垂体系统有复杂的相互作用。不同种的鼠达到性成熟的年龄各不相同,如鼢鼠要经过1年后才能达到性成熟,而田鼠则可在2个月达到性成熟。常年繁殖的动物,如家鼠,初情期多在体重达到成年动物的40%~60%或生长速度下降的初期。季节性繁殖动物的初情期受出生期的影响很大。
啮齿类动物的繁殖受光照、温度和食物来源等环境因素的影响,一般都在最适宜妊娠和幼子生活的植物生长季节繁殖;而家鼠由于环境因素和食物来源比较稳定,经过长期进化,其繁殖季节逐渐延长,甚至可以全年繁殖。啮齿类动物达到性成熟后,雌鼠会在一年繁殖期中有规律地多次动情,而雄鼠则会在整个繁殖期不断形成精子,因而出现生殖季节,如家鼠、家兔等。常年繁殖并不意味全年的繁殖活动毫无变化。大多数常年繁殖鼠种的生殖活动,在不同季节表现出有规律的高峰期和低潮期。
典型的季节性繁殖啮齿类动物每年只出现一个或两个繁殖季节。在繁殖季节内,雌鼠能一次或多次动情,雄鼠能不断形成精子。但在休情期内,卵巢或睾丸都不同程度地萎缩,甚至退化到幼鼠那样的程度,两性配子的形成完全停止。在休情期内,睾丸常缩回腹腔,各种依赖性激素控制的附性器官也都退化。
性周期,雌性动物在性成熟后,卵巢在神经和体液的调节下,出现周期性的卵泡成熟期和排卵期伴随着每次卵泡的成熟和排卵,整个机体特别是生殖器官,发生系列的形态和机能的变化,同时动物还出现周期性的性反射和性行为过程,称为动情周期( estrouscycle)或性周期( sexual cycle)。这种周期性的性活动过程除了妊娠期之外,一直延续到性机能停止的年龄为止。3种鼠类的动情期见表2-4。
动情周期是一系列逐渐变化的复杂生理过程,难以严格地加以区分,通常将其分为动情前期、动情期、动情后期和间情期。
动情前期( proestms)是性周期的准备阶段和性活动开始的时期。这时动物处于安静状态,没有交配欲的表现,但生殖器官却发生一系列变化,如卵巢内卵泡迅速生长,并达到成熟阶段;卵泡液体积增加,雌激素分泌并释放到血液中,刺激输卵管内壁的细胞生长,纤毛增多;子宫角蠕动加强,子宫内膜的血管分布大量增生;阴道上皮细胞增生加厚,整个生殖道腺体活动力增强。
动情期( estrus)是性周期的高潮期。这时动物出现兴奋不安,食欲减退,时常鸣叫和爬跨其他个体,并有交配欲。生殖器官的变化是:大多数动物出现卵巢中的成熟卵泡破裂和排卵,子宫黏膜血管大量增生、腺体分泌、子宫颈口张开、出现子宫蠕动和水肿,输卵管岀现蠕动,阴唇黏膜肿胀,有些啮齿动物从阴道流岀黏液。动情期啮齿类动物的各种表现和生殖器官的形态和机能的变化,其主要原因是由于卵泡所分泌的雌激素的作用所产生的。在一般情况下,啮齿类动物动情的同时往往引起排卵,但是,动情和排卵两种生理活动过程是可以分离而单独存在的。例如,切除卵巢的啮齿类动物,给予外源性雌激素仍能引起动情,但不能排卵,在生产实践中有些患有不孕的啮齿类动物,其原因之一是只有动情而不排卵。
动情后期( metaestrus)指动情结束后的一段时期。此期内雌性啮齿类动物恢复安静并拒绝交配。卵巢中形成了黄体,并分泌孕激素和雌激素。在孕激素的作用下,子宫为接受胚泡和适应胚泡的营养及附植发生一系列的变化。如果排出的卵细胞受精,动情周期就此中止,并开始进入妊娠阶段,直到分娩后才重新出现新的动情周期。如果排出的卵细胞没有受精,就过渡到间情期。
间情期( diestrus)是相对生理静止期。卵巢内的黄体开始退化,卵泡还没有开始发育。生殖道朝着恢复动情前期以前的状态退化,子宫内膜变薄,阴道上皮不角化等。
啮齿类动物的动情周期经常受到内外环境因素、营养及健康状况的影响。突然而剧烈的环境变化会造成动情周期的紊乱甚至停止。若以卵巢周期性变化特征来区分,可将性周期分为卵泡期( follicular phase)和黄体期( luteal phase),而排卵则是卵泡期和黄体期的分界线。卵巢周期和动情周期的关系为:
性周期的变化经常受内外环境因素、营养及健康状况的影响,主要受神经体液调节,其中激素的调节是重要基础。外环境变化的信息以及雄性刺激由雌性啮齿类动物的嗅觉、视觉、触觉感受器所接受,经过不同途径,通过神经系统影响到下丘脑一腺垂体一卵巢轴。性周期的几个阶段实质上是雌激素与下丘脑的GnRH、腺垂体的FSH、LH的分泌活动的相互协调。
卵泡初期(即动情之初期)是雌激素积累时期。此时雌激素浓度较低,对FSH、LH的分泌具有负反馈作用,但作用十分弱;此时卵泡靠内部的自我驱动机制加快了颗粒细胞分裂、增殖,雌激素分泌功能增强。
卵泡中期(动情前期的中、后期)重要的是卵泡及血液中雌激素浓度继续上升,并出现高峰。此时,卵母细胞已进入第1次成熟分裂早期,尽管有雌激素和卵泡抑制素的抑制性负反馈作用,但由于卵泡内的糖蛋白雄激素局部正反馈作用,通过卵泡膜细胞LH受体数量的增加,最终到达雌激素分泌高峰。
卵泡后期(进入动情期,排卵前夕)LH高峰出现。这是由于雌激素高峰的出现,对下丘脑、腺垂体产生正反馈作用,GnRH脉冲式分泌量及频率都增加,腺垂体对GnRH的敏感性也增加,因而出现LH分泌高峰,LH通过刺激孕激素分泌,影响到卵内酶的活性,而触发排卵。
黄体期(动情后期)LH刺激黄体细胞分泌孕激素、雌激素,后者对下丘脑和腺垂体产生负反馈作用,使GnRH分泌下降。同时当雌激素分泌量升高时,降低了下丘脑催乳素抑制激素的释放,而引起垂体前叶催乳素释放量增加,催乳素与LH协同作用,促进和维持黄体分泌孕酮。当孕酮分泌达到一定量时,对下丘脑和腺垂体也会产生负反馈作用,抑制垂体前叶FSH的分泌,以至卵巢卵泡不再发育,抑制宠物中枢神经系统的性中枢,使雌性啮齿类动物不再表现动情。同时孕酮也作用于生殖道及子宫,使之发生有利于胚胎附植的生理变化。如果排出的卵子已受精,囊胚刺激子宫内膜形成胎盘。此时黄体则继续存在下去成为妊娠黄体;若未受精,因没有取代的促性腺激素,黄体退化,雌激素、孕激素分泌下降并终止,对下丘脑-腺垂体的负反馈作用终止,又开始下一轮的周期。
松果体分泌的褪黑素( melatonin,MT)对下丘脑一垂体一性腺轴的功能活动有明显的抑制作用。切除幼年啮齿类动物的松果体,出现性早熟,性腺的重量增加,功能活动增强,因此认为松果体在青春期有抗性腺的功能。MT可抑制下丘脑GnRH的释放和腺垂体FSH和LH的分泌,同时也可以直接抑制性腺的活动,从而影响性周期。
4.排卵与排卵后黄体
(1)排卵过程
成熟卵泡壁发生破裂,卵细胞、透明带与放射冠随同卵泡液冲出卵泡,称为排卵( ovulation)。排卵过程从卵巢表面上皮细胞的逐步溶解开始,随后细胞开始退行性变化,并释放蛋白酶进入细胞间隙,使白膜、结缔组织、卵泡外膜和内膜崩解,整个卵泡壁也发生普遍性溶解。当破口处的颗粒细胞变性破坏时,卵细胞及其放射冠随同卵泡液被排人输卵管伞或腹腔。排卵前,卵泡经历着3大变化:①卵母细胞细胞质和细胞核成熟;②卵细胞聚合力松解,颗粒细胞各自分离;③卵泡膜变薄、破裂。
(2) 排卵机制
垂体脉冲式地释放大量LH是激发排卵的必要条件,而LH的释放又受到雌激素的正反馈调控。升高的雌激素在下丘脑经2-羟化酶催化,生成儿茶酚雌醇或儿茶酚雌酮,统称儿茶酚雌激素( catecholestron),这类激素具有儿茶酚和雌激素的双重作用,一方面能减少去甲肾上腺素(NE)的降解,并增强NE对下丘脑分泌Litre的刺激作用;另一方面也能在下丘脑和腺垂体内与雌激素竞争受体,抑制雌激素对下丘脑和垂体的负反馈调节。升高的雌激素一方面刺激下丘脑GnRH脉冲式分泌频率和幅度极大地增加;另一方面也使腺垂体对GnRH的反应超常的大,使LH分泌及在血液中的水平达到顶峰。LH通过刺激孕激素的生成,增强卵泡中许多蛋白水解酶的活性。LH也直接促进卵泡大量生成蛋白水解酶。蛋白水解酶活性的增强,是卵泡破裂的主要因素。此外,孕酮、前列腺素等激素也对排卵起重要作用。它们能增强纤维蛋白分解酶原的产生,从而增加纤维蛋白分解酶的活性,使卵泡膜分解破裂;并可刺激颗粒细胞合成孕酮及卵泡黄体化。则能对溶酶体膜起破坏作用,因此,可促进卵泡顶端上皮细胞溶酶体的破裂,使上皮细胞脱落,顶端形成排卵点,所有这些都为排卵创造条件。
(3)黄体的生成和退化
成熟卵泡破裂排卵后,由于卵泡液排出,卵泡壁塌陷皱缩,从破裂的卵泡壁血管流出血液和淋巴液,并聚积于卵泡腔内形成血凝块。此后颗粒细胞增生肥大,吸收类脂物质黄素,而变成黄体细胞,构成黄体的主体部分。同时卵泡内膜分生出血管,布满于发育中的黄体。在动情周期中,雌性啮齿动物如果没有妊娠,所形成的黄体在黄体期末退化,这种黄体称为周期性黄体。黄体退化时,颗粒细胞转化的黄体细胞退化很快,体积逐渐变小,细胞的数目也显著减少,颗粒层细胞逐渐被纤维细胞所代替,黄体细胞间结缔组织侵入、增殖,最后整个黄体颜色变白称为白体( corpus albicans),残留在卵巢内。大多数啮齿类动物的白体存在到下一周期的黄体期,即此时的功能性新黄体与大部分退化的白体共存。
上一篇:啮齿类动物的雄性生殖生理
下一篇:啮齿类动物受精与妊娠


